Медоносные пчелы вида Apis mellifera L. имеют естественный ареал распространения, охватывающий всю Африку, Европу, и Ближний Восток. В таком широком диапазоне они подразделились на 24 подвида, которые были сгруппированы вначале в три (Ruttner et al., 1978), а потом в четыре (Ruttner 1988) эволюционные ветви. На основе морфологической изменчивости медоносной пчелы F.Ruttner et al. (1978) предложили следующие три ветви: ветвь A, которая включала подвиды медоносной пчелы Африки (A.m.lamarckii, A.m.yemenitica, A.m.litorea, A.m.scutellata, A.m.adansonii, A.m.monticola, A.m.capensis, и A.m.unicolor); ветвь М., которая включала подвиды медоносной пчелы Северной Европы (A.m.mellifera), Португалии и Испании (A.m.iberiensis), и северной Африки (A.m.intermissa, и A.m.saharensis); ветвь C, которая включала подвиды медоносной пчелы Восточной Европы, Северного Средиземноморья, и Ближнего Востока.
Позже, F.Ruttner (1988) разделил эволюционную ветвь C в две группы, где подвиды A.m.sicula, A.m.carnica, A.m.ligustica, A.m.cecropia и A.m.macedonica были отнесены к эволюционной ветви C, в то время как другие подвиды (A.m.anatolica, A.m.adami, A.m.cypria, A.m.syriaca, A.m.meda, A.m.caucasica, и A.m.armeniaca), были отнесены в новую эволюционную ветвь O.
Дальнейшее подтверждение результатов классификации F.Ruttner et al. (1978) было обеспечено данными анализа микросателлитных локусов (Estoup et al., 1995; Franck et al., 1998; 2000) и митохондриальной ДНК (Smith et al., 1991; Garnery et al., 1992, 1993, 1995; Arias, Sheppard, 1996; Franck et al., 1998, 2000, 2001). По результатам анализа митохондриальной ДНК медоносной пчелы возникли некоторые разногласия с морфометрическими данными F.Ruttner (1988), где подвиды A.m.intermissa, A.m.saharensis (Garnery et al., 1992), и некоторые образцы A.m.iberiensis были включены в эволюционную ветвь А (Smith et al., 1991), а подвид A.m.lamarckii - в ветвь O (Franck et al., 2000b). W.S.Sheppard и M.D.Meixner (2003) в Центральной Азии обнаружили 25-й подвид медоносной пчелы, который был предположительно отнесен к ближневосточной эволюционной ветви О (Ильясов и др., 2006).
Вид Apis mellifera, благодаря широкому ареалу естественного распространения от Южной Африки до Северной Европы и от Центральной Азии до Западной Европы, где меняются климат по величине и продолжительности температуры, высоте над уровнем моря, уровню влажности, видовой состав растений-медоносов, разнообразие болезней и вредителей, приобрел очень высокие уровни изменчивости и генетического разнообразия. Большое разнообразие условий обитания ведет к образованию популяций медоносной пчелы с разными особенностями биологии, физиологии и поведения. Благодаря такой широкой экологической пластичности, вид Apis mellifera приобрел уникальную устойчивость и приспособленность к условиям окружающей среды. Существование таких экологически подразделенных популяций медоносной пчелы в течение нескольких миллионов лет привело к образованию подвидов. На сегодняшний день уровень подразделенности популяций находится на уровне подвидов. Вмешательство человека в жизнь пчел очень остро сказывается на их эволюции – человек, благодаря плановым и неконтролируемым внутривидовым скрещиваниям подвидов пчел практически остановил микроэволюционный процесс. Такое длительное воздействие на популяцию медоносной пчелы может привести к потере генофондов аборигенных подвидов, ареалы которых займут плохо приспособленные к окружающей среде гибридные популяции.
Для жителей Северной Евразии очень ценен для экономически выгодного разведения подвид Apis mellifera mellifera L., идеально приспособленный к жизни в холодном континентальном климате с продолжительными периодами с отрицательной температурой. Ни один другой подвид не способен к существованию в подобных климатических условиях без помощи человека. По F.Ruttner et al. (1978), F.Ruttner (1988), M.C.Arias, W.S.Sheppard (1996), А.В.Петухов и др. (1996), А.Г.Николенко и А.В.Поскряков (2002), Р.А.Ильясов и др. (2006, 2007) подвид медоносной пчелы A.m.mellifera имеет естественный ареал вдоль северной границы Евразии до примерно 600 с.ш.. Предполагается, что на такой обширной территории, протянутой вдоль Северной Евразии, не может обитать только один подвид медоносной пчелы A.m.mellifera с абсолютно идентичным геномом во всех локальных популяциях, а скорее всего существуют экологические группы или экотипы, различающиеся друг от друга рядом поведенческих, физиологических и морфологических признаков на уровне ниже подвидового. Некоторые называют пчелу этого подвида суперпородой (Кузьмичев, Гришин, 2008). Этот подвид медоносной пчелы в разных местах обитания имеет свое название и считается уникальным: в европейских странах ее называют темной европейской пчелой, в России – среднерусской пчелой. В республике Башкортостан среднерусскую пчелу называют башкирской пчелой, а башкирскую пчелу, обитающую, в диком виде в бортях называют бурзянкой, по названию Бурзянского района, где расположен заповедник Шульган-Таш, охраняющий этих пчел.
В России, как и в Европе, под воздействием антропогенного фактора произошла гибридизация A.m.mellifera с подвидами медоносной пчелы, распространенными в более южных широтах. В Европе произошла гибридизация A.m.mellifera в основном с подвидами A.m.carnica и A.m.ligustica, в России – с подвидами A.m.carnica, A.m.ligustica, A.m.caucasica. Проблема сохранения генофонда аборигенного подвида Северной Евразии решается уже давно. К сожалению, в Европе и России не удалось сохранить популяцию A.m.mellifera в границах естественного ареала – в большинстве разводят медоносную пчелу гибридного происхождения. Для сохранения генофонда аборигенных пчел необходима точная идентификация подвидовой принадлежности и происхождения семей. Первоначальные исследования, направленные на различение подвидов медоносной пчелы, были основаны на морфометрических (Goetze, 1940; Алпатов, 1948; Ruttner et al., 1978; Кривцов и др., 1995) и биохимических (Mestriner, 1969; Contel et al., 1977; Badino et al., 1982; Cornuet, 1983; Sheppard, Berlocher, 1984, 1985; Sheppard, McPheron, 1986; Sheppard, 1988; Del Lama et al., 1988, 1990; Lobo et al., 1989; Meixner et al., 1994; Ivanova et al. 2004; Simuth et al., 2004) методах изучения полиморфизма. В условиях все увеличивающегося уровня гибридизации эти методы начали терять свою эффективность.
Использование молекулярно-генетических методов оказалось более эффективным для различения подвидов медоносной пчелы с большим уровнем достоверности в условиях их массовой гибридизации. Начало генетических исследований медоносной пчелы было основано на анализе рестрикционного полиморфизма фрагментов (RFLP, ПДРФ) митохондриальной ДНК (mtDNA, мтДНК) (Hall, Smith,1991; Smith,1991; Garnery et al., 1992; Oldroyd et al., 1992; Schiff et al., 1995; Sihanuntavong et al., 1999). Дальнейшее развитие методов идентификации подвидов было основано на полимеразной цепной реакции (PCR, ПЦР). Первоначально в исследованиях генома медоносной пчелы нашел применение метод изучения полиморфизма со случайно выбранными праймерами (RAPD) (Kesseli, 1992; Greg, 1995; Zhanao, 1997; Чудинов, 1999; Sathees, 2001; Ivanova et al. 2004; Ivgin et al. 2004). Локус-специфичная ПЦР по микросателлитным локусам ядерной ДНК (Tares et al., 1993; Estoup et al., 1995; Rowe et al., 1997; Haberl, Tautz, 1999; Polazhek et al., 2000; Oldroyd et al., 2000; Sittipraneed et al., 2001; De La Rua et al., 2002, 2003; Solignac et al., 2003; Franck et al., 2000, 2001; Paar et al., 2004), по локусам кодирующих генов ядерной ДНК (Danforth et al., 2006; Shultz, Regier, 2000), по локусам митохондриальной ДНК (Cornuet et al., 1991; Hall, Smith, 1991; Crozier, Crozier, 1993; Moritz et al., 1994; Estoup et al., 1995; Lobo, 2000; Franck et al., 1998, 2000, 2001; Николенко, Поскряков, 2002; Ильясов и др. 2006, 2007) стала более точным методом для идентификации подвидов медоносной пчелы. Для идентификации подвидов медоносной пчелы также эффективно используется секвенционный анализ локусов ядерной и митохондриальной ДНК и анализ однонуклеотидных замен (SNP) (Itenov et al., 1991; Arias, Sheppard, 1996, 2005; Whitfield et al., 2006).
Нами были отобраны медоносные пчелы Apis mellifera из 43 семей с 2 пасек заповедника Шульган-Таш Бурзянского района республики Башкортостан (пасека Капова-Пещера кордон заповедника; пасека п.Коран-Елга), из 52 семей с 6 пасек Башкирской опытной станции пчеловодства Иглинского района республики Башкортостан (п.Улу-Теляк, пасеки Гареева, матковыводная, Орловская, Кугейко, Громова, Шамсуринская), из 37 семей с 8 пасек Юсьвенского района Коми-Премяцкого округа Пермского края (с.Архангельское, пасека Кривощекова Д.Ф.; с.Юсьва, пасека Быкова Н.А.; с.Почашор, пасека Сторожева В.М.; д.Федотово, пасека Власова В.Д.; д.Федорово, пасека Бояндина А.Г.; пасека д.Б.Они; пасека д.Доег; пасека д.Пожва). Всего были проанализированы пчел из 132 семей.
Для изучения генетической структуры населения медоносной пчелы Среднего и Южного Урала мы анализировали полиморфизм локуса COI-COII митохондриальной ДНК и микросателлитных локусов ap243, 4a110, a24 и a8. В трех локальных популяциях медоносной пчелы (бурзянская, иглинская и юсьвенская) наблюдались следующие частоты аллей (табл.1).
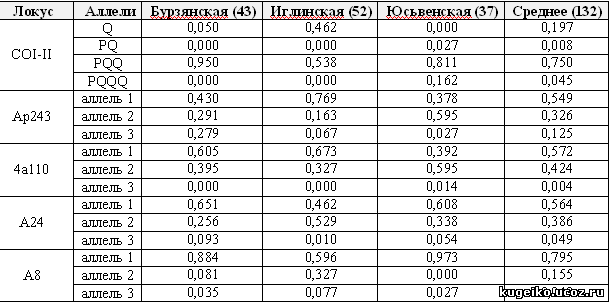
Таблица 1. Частота аллелей локуса COI-COII митохондриальной ДНК и микросателлитных локусов ap243, 4a110, a24 и a8 у исследуемых популяций пчелы медоносной
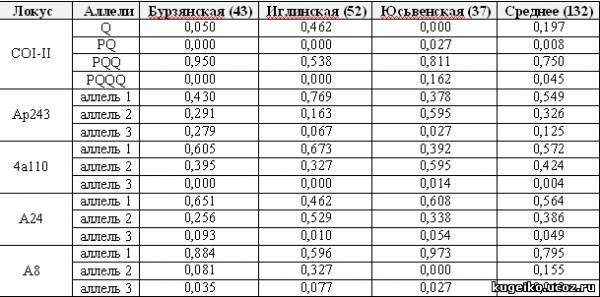 Межгенный локус COI-COII мтДНК у медоносной пчелы представлен 4 состояниями, где единственный элемент Q на Урале характеризует происхождение по материнской линии от подвидов пчел из южных регионов – A.m.caucasica, A.m.carnica, A.m.ligustica и др. Все остальные комбинации элементов P и Q на Урале характеризуют медоносную пчелу подвида A.m.mellifera по материнской линии. Как показывает табл.1, бурзянская и юсьвенская популяции характеризуются максимальным содержанием пчел, происходящих от A.m.mellifera по материнской линии.
Межгенный локус COI-COII мтДНК у медоносной пчелы представлен 4 состояниями, где единственный элемент Q на Урале характеризует происхождение по материнской линии от подвидов пчел из южных регионов – A.m.caucasica, A.m.carnica, A.m.ligustica и др. Все остальные комбинации элементов P и Q на Урале характеризуют медоносную пчелу подвида A.m.mellifera по материнской линии. Как показывает табл.1, бурзянская и юсьвенская популяции характеризуются максимальным содержанием пчел, происходящих от A.m.mellifera по материнской линии. Уровень гетерозиготности является одним из основных показателей в популяционно-генетическом анализе пчел, характеризующий величину биологического и генного разнообразия. Для популяции медоносной пчелы известно, что избыточная и недостаточная гетерозиготность ведет к неблагоприятным явлениям. Для популяции медоносной пчелы в норме, по результатам наших многолетних исследований в России, свойственен небольшой дефицит гетерозигот (Ильясов и др., 2008).
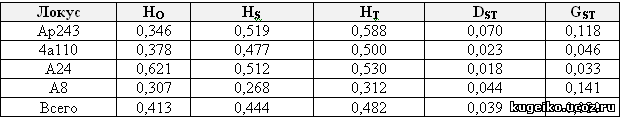
Мы рассчитали величину средней наблюдаемой гетерозиготности внутри субпопуляции HO (табл.2) и сравнили ее со средней ожидаемой гетерозиготностью внутри субпопуляции. Во всей популяции по всем микросателлитным локусам ядерной ДНК наблюдается дефицит гетерозигот. Гетерозиготность внутри субпопуляций НS больше гетерозиготности между субпопуляциями DST. Величина межпопуляционной генетической дифференциации GST для всей популяции небольшая и приближается к 0,1.
Показатели гетерозиготности для каждой локальной популяции также различались (табл.3). Наибольший дефицит гетерозигот наблюдался в иглинской локальной популяции, а наименьший - в бурзянской. Межпопуляционная составляющая гетерозиготности DST наименьшая в бурзянской локальной популяции, а наибольшая - в иглинской. Соответственно наименьшая величина межпопуляционной генетической дифференциации GST наблюдается в бурзянской локальной популяции, а наибольшая - в иглинской.
Таблица 2. Гетерозиготность во всей популяции по результатам анализа полиморфизма микросателлитных локусов ap243, 4a110, a24 и a8
HO – средняя наблюдаемая гетерозиготность внутри субпопуляции; НS – средняя ожидаемая гетерозиготность внутри субпопуляции; НT – средняя ожидаемая гетерозиготность в тотальной популяции; DST – средняя ожидаемая гетерозиготность между субпопуляциями; GST – величина межпопуляционной генетической дифференциации
Таблица 3. Гетерозиготность в локальных популяциях по результатам анализа полиморфизма микросателлитных локусов ap243, 4a110, a24 и a8
На основе изучения полиморфизма 4 микросателлитных локусов были рассчитаны коэффициенты инбридинга F-статистики для всей популяции (табл. 4). Расчет индекса фиксации (Wright, 1965) FIS, отражающего инбридинг особи относительно субпопуляции, показал, что во всей популяции наблюдается дефицит гетерозигот. Индекс фиксации FIT, отражающий инбридинг особи относительно всей общей популяции, также показал дефицит гетерозигот во всей популяции, но на более высоком уровне. Индекс фиксации FST, отражающий инбридинг между субпопуляциями относительно всей целой популяции, а также уровень дифференцированности популяции показал, что популяция в целом подразделена на субпопуляции.
Таблица 4. Коэффициенты инбридинга во всей популяции по результатам анализа полиморфизма микросателлитных локусов ap243, 4a110, a24 и a8
FIT – индекс фиксации, отражающий величину инбридинга на уровне всей популяции; FST – индекс фиксации, отражающий величину инбридинга между субпопуляциями и степень подразделенности популяции; FIS - индекс фиксации, отражающий величину инбридинга на уровне субпопуляций
Индексы фиксации, рассчитанные для субпопуляций были следующие (табл. 5). По значениям FST наименее подразделенной оказалась бурзянская популяция, а наиболее подразделенной иглинская. Наибольший уровень инбридинга и дефицит гетерозигот на уровне субпопуляций FIS и на уровне всей популяции FIT наблюдался в бурзянской популяции, а наименьший - в юсьвенской популяции. В иглинской популяции наблюдался аутбридинг с избытком гетерозигот.
Таблица 5. Коэффициенты инбридинга в локальных популяциях по результатам анализа полиморфизма микросателлитных локусов ap243, 4a110, a24 и a8
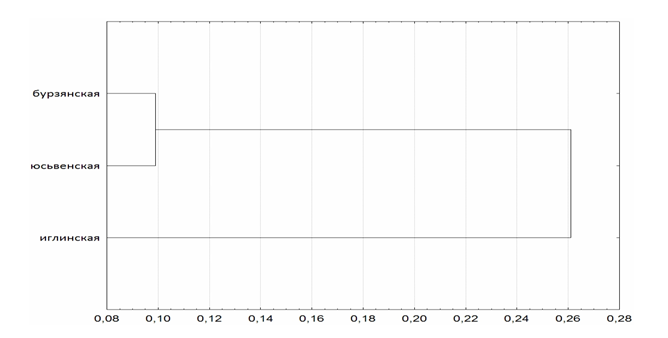
На основе стандартных генетических расстояний (Nei, 1978) по данным анализа полиморфизма локуса COI-COII митохондриальной ДНК и микросателлитных локусов ap243, 4a110, a24 и a8, была построена дендрограмма с использованием кластерного анализа методом ближайшего соседа, используя в качестве меры связи метод Варда, а меры расстояния - Евклидову дистанцию. На дендрограмме наблюдается распределение локальных популяций на два кластера, где максимально гибридизованная иглинская популяция группируется отдельно от двух других. Сходная картина распределения локальных популяций наблюдается и в случае анализа только микросателлитных локусов ядерной ДНК.
Рисунок. Дендрограмма построенная по результатам анализа полиморфизма локуса COI-COII митохондриальной ДНК и микросателлитных локусов ap243, 4a110, a24 и a8
Проведенный нами анализ генетической структуры трех удаленных друг от друга локальных популяций медоносной пчелы Южного и Среднего Урала показал их генетическую дифференциацию и подразделенность на субпопуляции в целом. Отмечалась совместная группировка юсьвенской популяции медоносной пчелы с бурзянской, которая ранее была определена на основе генетических исследований как популяция среднерусской пчелы A.m.mellifera. Это свидетельствует о происхождении юсьвенской локальной популяции медоносной пчелы по ядерной и митохондриальной ДНК от подвида A.m.mellifera. Ранее определенная на основе генетических исследований гибридная иглинская локальная популяция медоносной пчелы в данном исследовании располагалась отдельно. Следует отметить характерный для большинства популяций медоносной пчелы на Урале небольшой дефицит гетерозигот и инбридинг. Гибридная иглинская популяция, наоборот, характеризуется избытком гетерозигот и аутбридингом. Таким образом, на Южном (Бурзянский район республики Башкортостан) и Среднем Урале (Юсьвенский район Коми-Пермяцкого округа Пермского края) еще сохранились локальные популяции медоносной пчелы подвида A.m.mellifera с достаточными для длительного существования популяции во времени величинами ареала и численности.
Ильясов Р.А., Петухов А.В., Поскряков А.В., Николенко А.Г.
Институт биохимии и генетики Уфимского научного центра РАН
Материалы международной конференции ПЧЕЛОВОДСТВО - ХХI ВЕК. Пчеловодство, апитерапия и качество жизни. Москва 17 - 20 мая 2010 года. Страницы 72 - 77 |